Kirsten Kierulf-Vieira. LiS1, Båtsfjord kommune
Erlend Skaga. LiS1, Sykehuset Østfold
Einar Osland Vik-Mo. Seksjonsoverlege, Hjernesvulstseksjonen /Ass. gruppeleder , Vilhelm Magnus-laboratoriet, Nevrokirurgisk avdeling, Oslo universitetssykehus
Glioblastom er den vanligste og mest aggressive formen for hjernekreft. Allerede ved diagnosetidspunktet befinner det seg kreftceller over hele hjernen. Kreftceller med stamcelle-lignende egenskaper utgjør en del av disse cellene, og disse er resistente mot vanlig behandling. En stor heterogenitet i kreftsvulstens genetiske oppbygning gjør det vanskelig å finne standardiserte behandlinger som er effektive for alle glioblastompasienter. På Vilhelm Magnus-laboratoriet ved Oslo universitetssykehus jobber vi med å utvikle individualisert behandling basert på særtrekk ved kreftsvulstene som vi og andre har kartlagt. Dette laboratoriet ble startet for snart 40 år siden av professor Iver A. Langmoen, og oppkalt etter grunnleggeren av norsk nevrokirurgi Vilhelm Magnus. Siden oppstarten har laboratoriet arbeidet med en rekke basalbiologiske forskningsområder nært knyttet til klinisk nevrokirurgi, og over 20 PhD-kandidater har gjort arbeidet sitt her.
En helhjernesykdom
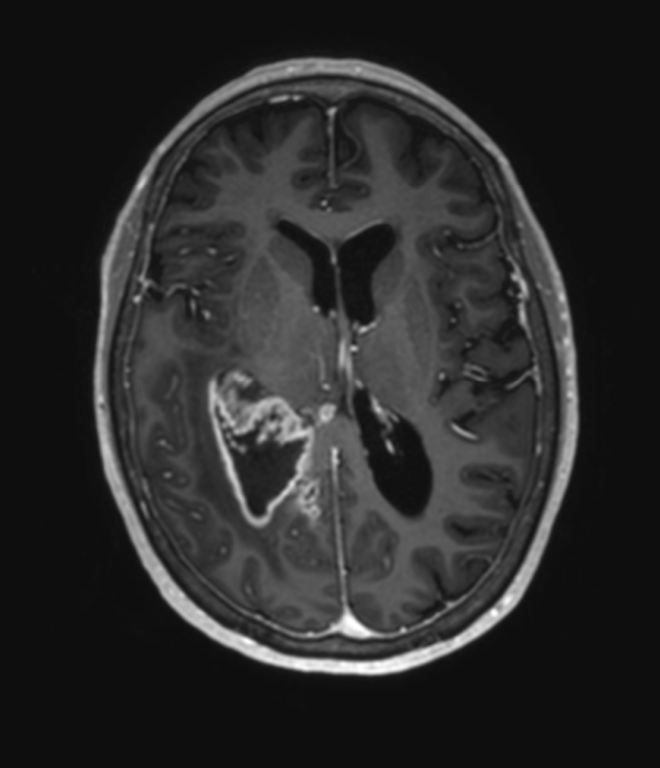
Figur 1. Glioblastom i høyre temporallappsregion. MR-bilde i T1-vektet sekvens med kontrast som viser utseende av et glioblastom med en kontrastladende svulst med sentralt henfall. Copyright forfatterne.
Til tross for intensiv behandling med maksimal sikker kirurgisk reseksjon, stråleterapi og kjemoterapi får de fleste glioblastompasienter residiv av svulsten i reseksjonsranden innen noen måneder. I en uselektert pasientpopulasjon er median overlevelse på under ett år (1). Når man vet at tumor kommer igjen på samme sted kan man spørre seg hvorfor man ikke fjerner mer hjernevev ved den primære kirurgien? Det tenkte nevrokirurgen Walter Dandy også på 1920-tallet, og gjennomførte hemisfærektomier som den kirurgiske behandlingen av glioblastom. Imidlertid fikk pasientene da residiv i motsatt hjernehalvdel (2). Dette forteller oss at pasienten ved diagnosetidspunktet har kreftceller spredt over hele hjernen.
Kirurgi er imidlertid viktig og god kreftbehandling – det er bare ikke tilstrekkelig. Kirurgi ble introdusert som rutinemessig behandling i første halvdel av 1900-tallet da reseksjon av tumor ble vist å øke median overlevelse fra ca. tre til syv måneder. (3). Kirurgi er også helt sentralt for å sikre tumorvev til patologisk vurdering for etablering av korrekt diagnose. Det neste behandlingsalternativet som viste en bedret overlevelse for glioblastom kom på 1960-tallet med stråleterapi. Å behandle glioblastom med både kirurgi og stråling viste da en økt overlevelse på ytterligere ca. fem måneder (4). Det skulle så gå flere tiår før man i 2005 viste at det kjemoterapeutiske middelet temozolomide ga en signifikant bedring av median overlevelse med omtrent tre måneder, gitt som tilleggsbehandling til kirurgi og stråling (5). Til tross for disse fremskrittene står fortsatt glioblastom igjen som en av de raskest dødelige sykdommene innen onkologien.
Kreftstamceller driver kreftutviklingen
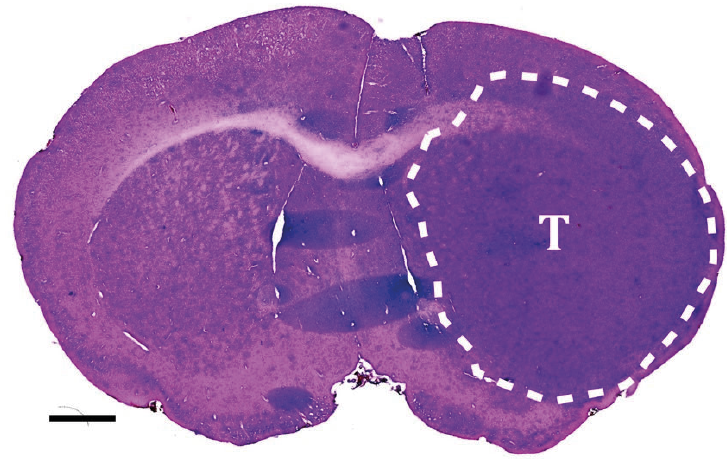
Figur 2. Ved å transplantere kreftstamceller til hjernen hos immundefekte mus danner kreftstamcellene en invasiv svulst med histopatologiske trekk som ved et glioblastom. Snittet er fra en musehjerne farget med hematoxylin & eosin. Svulsten er avgrenset makroanatomisk med den stiplede linjen og markert med bokstaven T. Skaleringslinje 1 mm. Copyright forfatterne.
Kreftstamcellehypotesen har røtter tilbake til 1858 da den tyske legen Rudolf Virchow postulerte at kreft består av umodne celler (6). De første solide bevisene fikk vi først i 1997 da Dick og kollegaer identifiserte en liten subpopulasjon av leukemiceller med egenskaper som stamceller. Ved transplantasjon til mus kunne disse cellene gi opphav til en kreftsykdom som lignet pasientens egen (7). Etter hvert ble det identifisert slike cellepopulasjoner i en rekke krefttyper, inklusive glioblastom (8). Som enhver ny teori møtte den også raskt motstand, og det ble blant annet vist at kreftstamceller kunne bli til ”vanlige” kreftceller og vice versa (9).
Til tross for at kreftstamcelleteorien har noen konseptuelle svakheter har den praktisk verdi. Kreftceller med stamcelletrekk er mer aggressive enn andre kreftceller, og når de transplanteres til mus gir de opphav til kreftsvulster som bevarer viktige særtrekk ved originalsvulsten (10-12). Vi kan dyrke pasientbiopsier i et medium som stimulerer til vekst i tredimensjonelle sfærer. I et slikt medium trives kreftstamcellene godt, mens de mindre potente kreftcellene har vanskeligere for å overleve. Dette gjør at vi kan selektere de cellene som er mest relevante for å forstå kreftens særegenheter, holde pasientens aggressive kreftstamceller i live på laboratoriet, og teste nye behandlingsmåter på dem. Dette gjør kreftstamcellene til en god modell for individualisert hjernekreftforskning.
Ingen pasienter er like
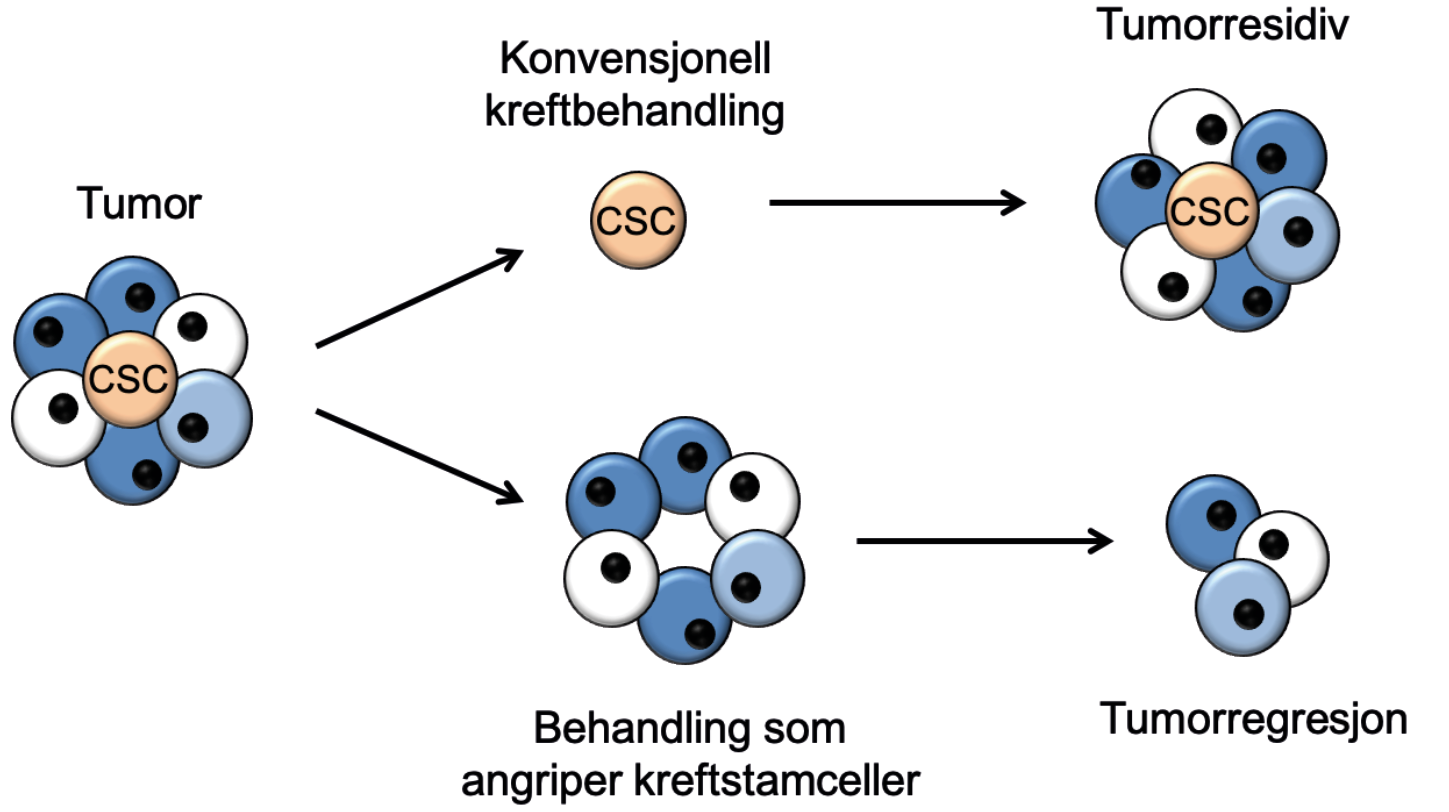
Figur 3. Skjematisk fremstilling av kreftstamcelleteorien. Denne postulerer at konvensjonelle kreftterapier angriper ”vanlige” kreftceller, men lar kreftstamcellene (CSC; cancer stem cells) være, og dermed tillater svulsten å vokse tilbake (øverst). Ved å bruke terapier rettet mot kreftstamcellene vil svulsten miste sin evne til selvfornyelse og til slutt forsvinne (nederst). Adaptert fra Reya et al. 2001. Copyright forfatterne.
I hjernekreft foreligger det i de ulike celletypene et hierarki av potensial for videre vekst av svulsten, der kreftstamcellen er på topp. Det danner et grunnlag for heterogenitet i hver enkelt svulst – intratumor-heterogenitet (10). Innen hjernekreft er det også en stor intertumor-heterogenitet, altså en variasjon i kreftsvulstens genetiske og cellulære oppbygning fra pasient til pasient. Det viste seg raskt at ulike pasienter har varierende effekt av behandling med temozolomide (13). Dette er et godt eksempel på klinisk relevant heterogenitet. Behandlingseffekten er avhengig av epigenetisk regulering av genet MGMT, som er et DNA-reparasjonsprotein. Dersom MGMT er skrudd på, er cellen resistent mot temozolomide-indusert celleskade fordi den reparerer skadet DNA fortløpende. Dersom MGMT er skrudd av, er cellen imidlertid mer sårbar for temozolomide-indusert celleskade, og behandlingen øker overlevelse (13). Har denne kunnskapen noen behandlingsmessig konsekvens per i dag? Foreløpig er svaret nei. Ettersom vi ikke har noen alternativ behandling å tilby, får alle pasientene tilbud om temozolomide.
Det er dermed flere utfordringer vi står overfor i behandlingen av en pasient med glioblastom:
- Det er en helhjernesykdom – komplett kirurgisk reseksjon av tumor er ikke mulig.
- Svulsten inneholder kreftstamceller med høy grad av resistens mot behandling.
- Potensielle behandlingsmetoder må krysse blod-hjerne-barrieren.
- Det er stor intertumor-heterogenitet, noe som gjør det vanskelig å finne en behandling som passer for alle.
Behandlingsstrategier
Enkeltgener/signalveier/mekanismer
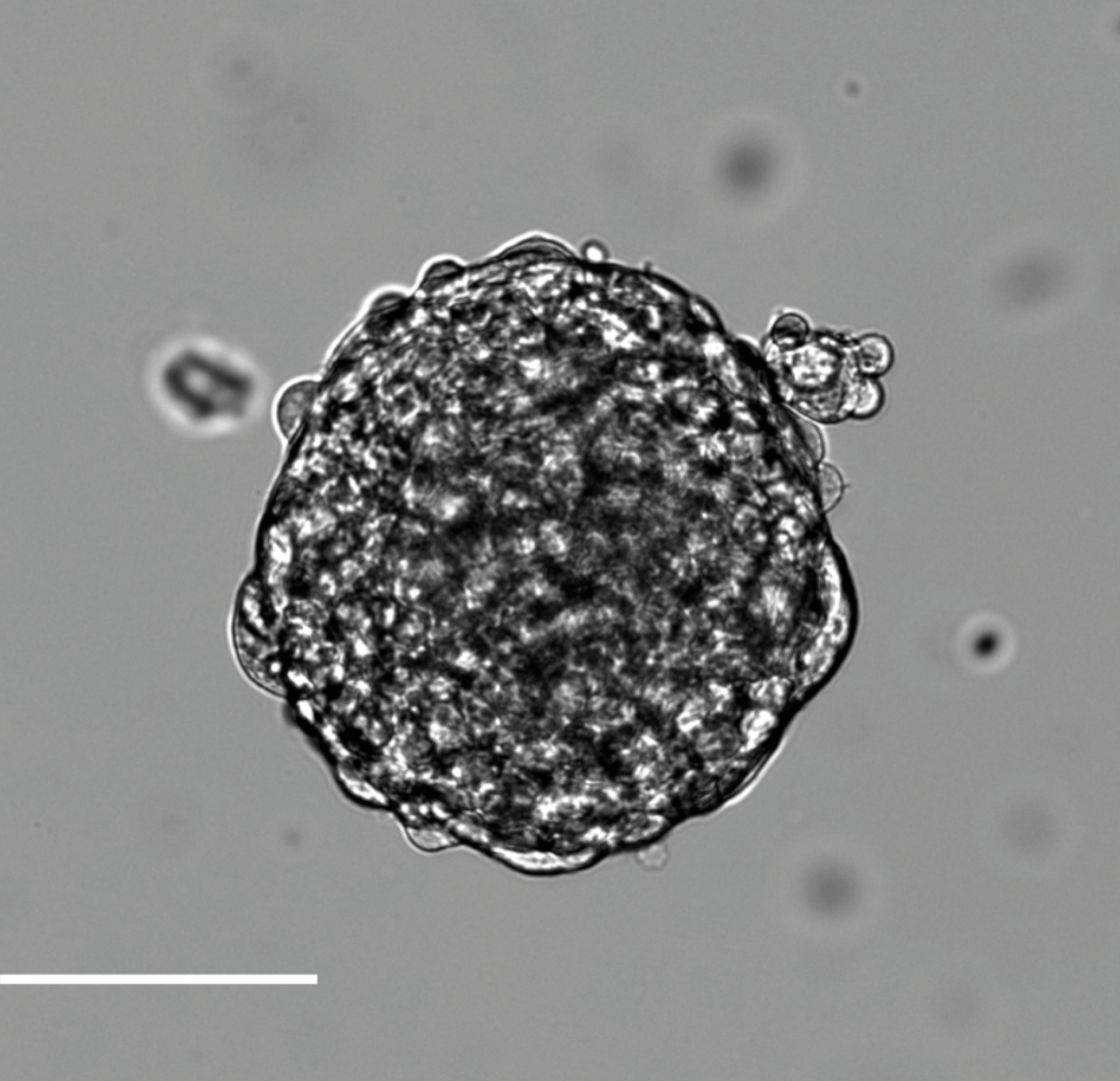
Figur 4. Ved å dyrke kreftstamceller fra glioblastom danner de tredimensjonale sfærer i cellekultur. Skaleringslinje 100 µm. Copyright forfatterne.
Vi har sammenlignet kreftstamceller fra glioblastom med normale hjernestamceller, og identifisert både enkeltgener og signalveier som var forskjellig uttrykt i de to celletypene. Et av funnene var at den intracellulære signaleringskaskaden WNT var oppregulert i kreftstamcellene fra glioblastom. Tidligere er WNT-signalering vist å bidra til kreftutvikling i blant annet tarmvev og i medulloblastom – en hjernekreft som forekommer oftest hos barn (14). Vi blokkerte WNT-signalet på to ulike måter og fant at dette fikk kreftstamcellene fra glioblastom til å dele seg > 50 % saktere, uten å påvirke de normale hjernestamcellene (15, 16). Denne effekten gikk via cellesyklus og apoptose (15), to grunnleggende celleprogrammer som alltid er endret i kreft. Utfordringene med å overføre slike resultater til klinikken er flere. Signalveiene som er endret ved kreftsykdommer er også til stede i friske celler, og innehar gjerne en viktig funksjon i disse cellene. Potensielle hemmere må også krysse blod-hjerne-barrieren som beskytter hjernen mot fremmede stoffer. En betydelig utfordring er også den store heterogeniteten i hvor sensitive kreftstamcellene fra ulike pasienter er for behandlingen.
Individualisert kjemoterapi
I Vilhelm Magnus-laboratoriet har vi over flere år dyrket og karakterisert kreftstamceller i pasientbiopsier fra glioblastom. Vi har observert en stor biologisk heterogenitet mellom pasienters kreftstamceller med henblikk på blant annet celledelingshastighet, differensieringspotensial in vitro samt vekstmønster og invasivitet in vivo (17). Den store heterogeniteten gjør at individualiserte cellekulturer med kreftstamceller representerer en forenklet, men levende modell av pasientens egen svulst med muligheter for eksperimentell utprøving og individualiserte behandlinger. Med inspirasjon fra infeksjonsmedisin og testing av bakteriers antibiotikasensitivitet har vi i samarbeid med Universitetet i Helsinki etablert en klinisk protokoll til en multisenterstudie for individualisert kreftbehandling med kjemoterapi: Vi dyrker pasientbiopsier ved Vilhelm Magnus-laboratoriet, deretter flys de til Helsinki og testes for følsomheten for over 450 ulike kreftlegemidler med automatisert teknologi. Det har avdekket en kompleks intertumor-heterogenitet i hvilke legemidler individuelle kreftstamceller er følsomme for, med stor variasjon innen alle typer klasser av kjemoterapeutiske legemidler, fra klassiske cellegifter til målrettede tyrosinkinasehemmere. På bakgrunn av resultatet planlegger vi å behandle pasienter med residiv av glioblastom etter en skreddersydd legemiddelprofil.
Kreftvaksine

Figur 5.WNT styrer uttrykket av viktige gener relatert til celledeling, apoptose og cellemotilitet. Signalveien aktiveres ved at WNT-proteinet binder seg til en reseptor på overflaten av cellen, som igjen fører til at proteinet b-catenin akkumulerer, translokerer til cellekjernen og aktiverer transkripsjon. Vi har vist at WNT er oppregulert i kreftstamceller fra glioblastom, og at ved å blokkere WNT-signalet kan vi redusere celledelingshastigheten med > 50 % uten å påvirke de normale hjernestamcellene. Copyright forfatterne.
I samarbeid med blant annet Seksjon for celleterapi og Seksjon for utprøvende kreftbehandling har vi brukt kreftstamceller og dendrittiske celler til å utvikle en individualisert terapeutisk kreftvaksine for glioblastompasienter i en fase I/II-studie. Først etablerte vi individualiserte cellekulturer med kreftstamceller fra tumorbiopsier, før vi renset og oppkopierte mRNA fra kreftstamcellene. Deretter høstet vi monocytter ved leukaferese og modnet dem til antigenpresenterende dendrittiske celler. Til slutt transfekterte vi det amplifiserte mRNA fra kreftstamceller, samt de to kjente tumorassosierte antigenene survivin og hTERT, til de modne dendrittiske cellene. Disse cellene kunne så oversette mRNA til protein og presentere dette på overflaten. De dendrittiske cellene ble injisert intradermalt hos pasienter med regelmessige intervaller. Selv om de fleste pasientene fikk tumorresidiv til slutt, så vi en forbedret overlevelse hos disse pasientene sammenlignet med historisk tilpassede kontroller. Tre av disse pasientene har levd leger enn syv år, og er fortsatt i live. Andelen av langtidsoverlevere er sammenlignbart med funn i andre immunterapistudier. Vi har nå startet en randomisert fase II/III-studie hvor vi totalt skal inkludere 60 pasienter til enten standardbehandling eller dendrittcelleterapi etter gjennomført stråle/kjemoterapi.
Veien videre
To av de viktigste lærdommene fra de siste tiårenes kreftforskning er at kreften har en avansert homeostase på linje med andre vev, og at hver kreftsykdom er unik. Individualisert kreftbehandling er et forsøk på å finne svakheter i kreftens virkemåte, der standardbehandling kommer til kort. Dette er spesielt tydelig i glioblastom, som er en heterogen kreftsykdom med svært dårlig prognose. Videre forskning vil derfor, i tillegg til å utvikle nye behandlingsmetoder, fokusere på å finne robuste og kostnadseffektive måter å predikere hvilken behandling pasienten vil ha nytte av.
«Individualiserte -cellekulturer med kreftstamceller en -levende modell med muligheter for eksperimentell utprøving og -individualisert -behandling»
Referanser
- Ronning PA, Helseth E, Meling TR, Johannesen TB. A population-based study on the effect of temozolomide in the treatment of glioblastoma multiforme. Neuro Oncol. 2012;14(9):1178-84.
- Dandy W. Removal of Right Cerebral Hemisphere for Certain Tumors with Hemiplegia. Preliminary Report. JAMA. 1928;90(11):823-5.
- Davis L, Martin J, et al. A study of 211 patients with verified glioblastoma multiforme. J Neurosurg. 1949;6(1):33-44.
- Kristiansen K, Hagen S, Kollevold T, Torvik A, Holme I, Nesbakken R, et al. Combined modality therapy of operated astrocytomas grade III and IV. Confirmation of the value of postoperative irradiation and lack of potentiation of bleomycin on survival time: a prospective multicenter trial of the Scandinavian Glioblastoma Study Group. Cancer. 1981;47(4):649-52.
- Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005;352(10):987-96.
- Virchov R. Die Cellulapathologie in ihrer Begrundung auf physiologische und pathologische Gewebelehre. Berlin: August Hirschwald. 1858.
- Bonnet D, Dick JE. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med. 1997;3(7):730-7.
- Ignatova TN, Kukekov VG, Laywell ED, Suslov ON, Vrionis FD, Steindler DA. Human cortical glial tumors contain neural stem-like cells expressing astroglial and neuronal markers in vitro. Glia. 2002;39(3):193-206.
- Chen R, Nishimura MC, Bumbaca SM, Kharbanda S, Forrest WF, Kasman IM, et al. A hierarchy of self-renewing tumor-initiating cell types in glioblastoma. Cancer Cell. 2010;17(4):362-75.
- Lan X, Jorg DJ, Cavalli FMG, Richards LM, Nguyen LV, Vanner RJ, et al. Fate mapping of human glioblastoma reveals an invariant stem cell hierarchy. Nature. 2017;549(7671):227-32.
- Rosenberg S, Verreault M, Schmitt C, Guegan J, Guehennec J, Levasseur C, et al. Multi-omics analysis of primary glioblastoma cell lines shows recapitulation of pivotal molecular features of parental tumors. Neuro Oncol. 2017;19(2):219-28.
- Davis B, Shen Y, Poon CC, Luchman HA, Stechishin OD, Pontifex CS, et al. Comparative genomic and genetic analysis of glioblastoma-derived brain tumor-initiating cells and their parent tumors. Neuro Oncol. 2016;18(3):350-60.
- Hegi ME, Diserens AC, Gorlia T, Hamou MF, de Tribolet N, Weller M, et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N Engl J Med. 2005;352(10):997-1003.
- Barker N, Clevers H. Mining the Wnt pathway for cancer therapeutics. Nat Rev Drug Discov. 2006;5(12):997-1014.
- Kierulf-Vieira KS, Sandberg CJ, Grieg Z, Gunther CC, Langmoen IA, Vik-Mo EO. Wnt inhibition is dysregulated in gliomas and its re-establishment inhibits proliferation and tumor sphere formation. Exp Cell Res. 2016;340(1):53-61.
- Sandberg CJ, Altschuler G, Jeong J, Stromme KK, Stangeland B, Murrell W, et al. Comparison of glioma stem cells to neural stem cells from the adult human brain identifies dysregulated Wnt- signaling and a fingerprint associated with clinical outcome. Exp Cell Res. 2013;319(14):2230-43.
- Fayzullin A, Sandberg CJ, Spreadbury M, Saberniak BM, Grieg Z, Skaga E, et al. Phenotypic and Expressional Heterogeneity in the Invasive Glioma Cells. Transl Oncol. 2019;12(1):122-33.